












【佳学基因检测】股骨头坏死的基因解码及其基因检测
为什么要做股骨头坏死的基因解码、基因检测?
股骨头坏死(FHON)是一个世界性的具有挑战性的临床课题。使用类固醇是股骨头坏死的主要病因之一。有几种与股骨头坏死相关的遗传变异。因此,股骨头坏死的基因解码基因检测的目的是提供对与非甾体和甾体诱导的股骨头坏死相关的基因突变的荟萃分析和系统评价的全面总结。
股骨头坏死的基因解码方法
符合条件的研究选自 PubMed 和 MEDLINE 数据库,用于收集各种系统的荟萃分析和评论。使用人类基因组流行病学网络的威尼斯标准分配遗传主效应评分,以评估单核苷酸多态性 (SNP) 对股骨头坏死影响的累积证据。
股骨头坏死基因解码的研究结果
八个研究报告了对基于 SNP 的候选研究的荟萃分析,涵盖 8 个基因的 13 个基因突变基因检测。在非类固醇诱导的股骨头坏死基因突变中,包括 MMP8 中的 jxjy2012390 和 jxjy11225394,肿瘤坏死因子 (TNF)-α 中的 jxjy1800629 和 jxjy361525,内含子 4 中的 VNTR,内皮一氧化氮合酶 (eNOS) 中的 jxjy1799983 和 jxjy2070744,血管内皮生长中的 jxjy2010963因子 (VEGF) 和因子 V 中的 jxjy6025 在每个研究中都显示出显著性。类固醇诱导的股骨头坏死基因突变基因检测包括载脂蛋白 (Apo)B 中的 jxjy693 和 jxjy1042031、ABCB1 中的 jxjy1045642 和 PAI-1 中的 jxjy1799889 在每个研究中都显示出显着性。
股骨头坏死的基因解码结论
根据股骨头坏死的基因解码研究中进行的系统评价,股骨头坏死的基因解码基因检测重大课题研究组组织了与股骨头坏死相关的基因组并检查了每一个位点对疾病发生的贡献。股骨头坏死的基因解码基因检测重大课题研究组的结果可以为理解股骨头坏死的致病机制提供一种综合的基因检不则方法。预计这些结果可能有助于预诊断策略,评估非类固醇诱导和类固醇诱导的股骨头坏死的个体风险。
股骨头坏死基因解码基因检测关键词
股骨头坏死,伞形检查,类固醇,多态性,基因突变,基因检测
人体基因序列变化与人体疾病特征关于股骨头坏死与基因检测意义的分析
股骨头坏死是一个世界性的具有挑战性的临床课题。法国解剖学家和病理学家 Jean Cruveilhier 将股骨头坏死描述为伴有晚期并发症的髋部创伤。从那时起,许多研究试图确定股骨头坏死的病因。根据对股骨头坏死进行的病理学研究,发病机制的分类包括以下内容。首先,据报道,由于血流受损导致特定区域的循环受阻是股骨头坏死的最终途径。其次,类固醇诱导的股骨头坏死发病机制不同,是一种骨内间隔综合征。第三,股骨头坏死病因复杂,包括遗传因素和受环境危险因素影响。这意味着遗传因素可能是疾病出现的原因,并且与其他发病因素密切相关。在存在这些遗传易感因素的情况下,股骨头坏死的风险会增加。
基因解码表明使用类固醇是股骨头坏死的主要病因之一。类固醇是股骨头坏死的剂量依赖性发病原因。长期使用类固醇可能导致股骨头塌陷、结构改变和髋关节功能障碍。类固醇诱发股骨头坏死的发病机制有多种学说,包括(1)类固醇通过增加皮下脂肪动员引起微血管内脂肪栓塞的脂质代谢紊乱学说;(2) 供血不足理论认为某些类固醇会干扰股骨头的血液供应并导致血管内血栓形成; (3) 骨髓基质细胞 (BMSCs) 的脂肪生成,这是参与类固醇诱导的股骨头坏死发生和进展的主要机制。尽管许多理论被认为是类固醇诱发股骨头坏死的病因,但确切的病理过程仍不清楚。
到目前为止,已经解码了几种与股骨头坏死相关的遗传因素,包括参与脂质代谢和凝血系统的载脂蛋白 (Apo)A;VEGF;因子 V Leiden (FVL)的突变;和纤溶酶原激活物抑制剂 1 型多态性。此外,还报道了与类固醇诱导的股骨头坏死相关的基因因素,包括与脂质转运相关的基因,如 ABCB1 和 CYP3A ;以及与血管闭塞和凝血障碍有关的遗传多态性,例如 PAI 和 MTHFR。这些基因信息因素是了解股骨头坏死的非常有用的工具。识别有和没有可以导致股骨头坏死的有关的特定的基因序列将为股骨头坏死的诊断和治疗提供极有价值的信息。
因此,股骨头坏死的基因解码基因检测的目的是对对与非甾体和甾体诱导的股骨头坏死相关的基因突变进行全面分析,以提供股骨头坏死相关的基因检测,并为诊断、治疗和早期风险干预提供基因检测服务。
股骨头坏死的基因解码分析方法
大数据搜索策略和研究数据选择评判标准
自 PubMed 和 MEDLINE 数据库中选择符合条件的研究,用于根据系统评价和 Meta 分析指南的首选报告项目收集各种系统性 Meta 分析和评价。搜索策略包括关键字“骨坏死”[MeSH 术语] OR “骨坏死”[所有字段] OR “avascular”[所有字段] AND “坏死”[所有字段] OR “avascular necrosis”[所有字段])和“基因"[MeSH 术语] OR "genes"[All Fields] OR "gene"[All Fields] AND "meta"[Journal] OR "meta"[All Fields]。检索到的出版物由两位基因解码分析师按照标准再进行独立筛选。根据共识解决分歧。如果无法做出最终决定,则请出第三名资深基因解码分析师。在筛选标题和摘要后,再对研究结果的全部内容进行深入评价。
符合条件的研究包括四种类型的荟萃分析:(i)基于 SNP 的系统评价和与股骨头坏死相关的基因荟萃分析,(ii)提供有关统计过程和结果的明确信息的研究,以及(iii)研究以英文撰写并于 2012 年 1 月 1 日之后发表。本次基因解码分析聚焦成人人群,并且不考虑性别差异,因为与股骨头坏死相关的基于 SNP 的一般荟萃分析不分性别进行。基因解码将以下类型的研究排除在外:(i) 没有对证据进行定量综合的系统性荟萃分析,(ii) 非人类研究,(iii) 存在巨大错误或质量差,以及 (iv) 没有特定统计数据如异质性检验、检查的风险因素和总体p价值的研究结果。
股骨头坏死如何提行研究数据的提取
数据提取阶段工作通过两次两次独立的分析进行。第一次是由基因解码师提取数据,第二步由另一位基因解码人员对数据进行检查。为选择符合条件的研究数据,所有研究结果应包括以下信息: (i) 第一作者姓名;(ii) 出版年份;(iii) 每项研究的基因突变基因检测和效应大小 (ES);(iv) 每篇文章中包含的研究数量;(v) 样本量、p值和异质性估计。
股骨头坏死的方法学的可信度分析
方法学可信度质量评价通过多重系统评价评估 (AMSTAR) 工具进行评级。该版本包括 16 个项目,用于评估研究结果的方法学的可信度。该评估按从高质量到非常低质量的等级进行评级。
统计分析
使用人类基因组流行病学网络的威尼斯标准分配遗传主效应分数,以评估 SNP 对股骨头坏死影响的累积证据。简而言之,这些指南提供了评估遗传流行病学累积证据的标准。证据的数量、复制的程度和防止偏倚是这些指南中的指标。三个级别的证据是强、中或弱。根据这些标准,大量证据和大样本量确保了检测关联的足够能力。通过对数千个多态性的全基因组测试,股骨头坏死的基因解码基因检测重大课题研究组用p评估了主要遗传效应评分类别的证据至少 < 0.005 的值被认为是强候选资格。表中提供了指南应用的具体描述表1.
表1:扩展人类基因组流行病学网络威尼斯标准以评估基因主效应的描述
| 考虑标准 | 类别 | 基因解码操作方案 |
|
证据数量 |
A | 样本量超过 1000 |
| B | 样本量 100–1000 | |
| C | 样本量小于 100 | |
|
重复 |
A | I2 < 50% |
| B | 25% < I2 < 50% | |
| C | I2 > 50% | |
|
偏差规避 |
A | 考虑偏差,例如基因分型偏差、人口分层和选择性报告偏差 |
| B | 基因型定义中的偏差=未报告所做的/没有质量控制检查/适当的质量控制检查 | |
| C |
人口分层 = 未报告做了什么/什么都没做/相同的血统组/针对报告血统的调整/基于家庭的设计/基因组控制、PCA 或类似方法
选择性报告偏差=已发表数据的元分析/回顾性努力以包括未发表的数据/联盟内的元分析 |
证据分为强、中和弱。当每个标准都获得 A 时,证据被归类为强。当在每个标准中都没有得分 C 但没有 AAA 时,证据被归类为中等。弱证据在三项标准中的一项中记录为 C
股骨头坏死的基因解码结果
确定的文章数量和类型
对于系统评价,从数据库中确定了 24 个研究。剔除不符合研究主题的研究后,剩下13个研究。另外三个研究被评为不合格的研究而被排除,因为它们提供的数据不足。排除了其他两个没有显着结果的研究。最后,选择了八个符合条件的研究数据,因为它们包括令人信服的数据、适当的定性评估、显着的结果、是对人的研究和符合股骨头坏死的基因解码基因检测重大课题研究组目的的主题(图 1)。 在 8个研究数据中,有 5 个非类固醇诱导的股骨头坏死的相关研究,包括 9 种基因突变; 3 个类固醇诱导的股骨头坏死相关研究,包括 4 种基因突变。所有研究都是回顾性研究,以确定股骨头坏死相关的基因突变基因检测。研究中包含的所有数据都使用血液样本对受试者特异性基因分型基因检测。
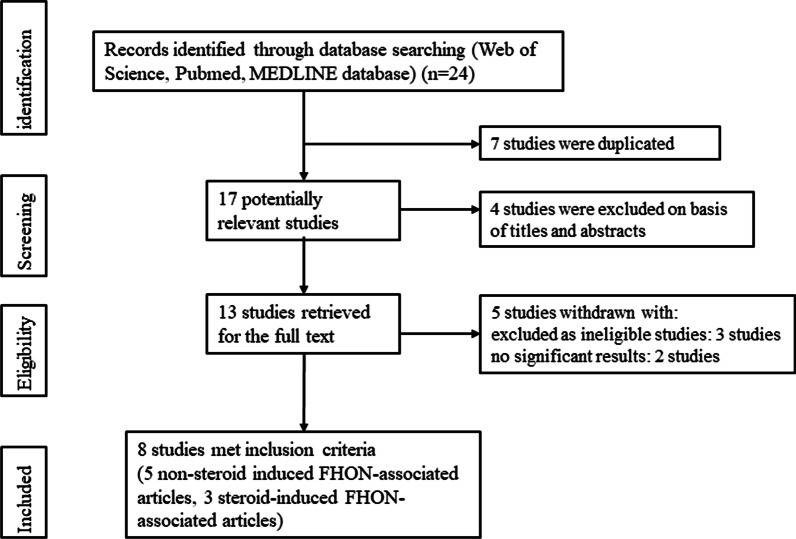
图1:研究选择过程的流程图
采用SNP基因检测所得到的主要结果
在国际范围内共有八个创新性研究得出了基于 SNP 的候选研究的荟萃分析,涵盖 8 个基因和 13 个基因突变。
在非类固醇诱导的股骨头坏死中,MMP-8 在 jxjy2012390 和 jxjy11225394 中显示出有意义的结果。jxjy11225394 非常显着,遗传主效应的p值低于 0.00001。根据威尼斯标准,jxjy11225394 SNP 显示出强有力的证据类别。彭等人证明了作为TNF-α的SNP的jxjy1800629和jxjy361525与G等位基因显示出低优势比。在 jxjy361525 中,等位基因遗传模型和隐性遗传模型的优势比低于 0.3,p值低于 0.001。jxjy361525 的这一结果显示了极其重要和强有力的证据,在威尼斯标准中得分为 AAB。宋和李等人报道了 eNOS 基因内含子 4 中可变数量的串联重复序列 (VNTR) 在等位基因遗传模型和显性遗传模型中显示出很高的显着性,p值低于 0.001,证据等级适中。jxjy1799983 和 jxjy2070744 也显示出几乎不显着的结果和过度的异质性,I 2值分别为 80.20% 和 90%。亚洲人的 VEGF 基因 jxjy2010963 在纯合子模型和显性模型中也显示出显着的结果,有强有力的证据。一个_在 jxjy6025 中记录了 < 0.001 的值,具有强有力的证据,这是因子的基因突变。尚等人表明混合种族人群中的 jxjy6025 显示疾病发生的可能性很高,优势比为 4.55(表2)。
表 2:来自非类固醇诱导的股骨头坏死的合格荟萃分析的基因突变基因检测
| 基因突变基因检测 | 基因(或近基因) | 参考(数据收集年份) | 种族 | 样本量(病例/对照) | 纳入研究 | 次要等位基因和参考等位基因 | 遗传方式 | 效应模型号类型 | 优势力比(95% CI) | 遗传主效应的p值 | 异质性 (%) | 威尼斯标准 | 证据等级 |
|
jxjy2012390 |
MMP-8 |
江等人 |
亚洲人 |
884/1012 |
3 |
A&G |
等位基因模型 | 固定效果 | 1.34 [1.00–1.79] | 0.005 | 0 | 不适用 | 不适用 |
| 杂合子模型 | 固定效果 | 1.40 [1.14–1.73] | 0.002 | 0 | 不适用 | 不适用 | |||||||
| 显性作用 | 固定效果 | 1.39 [1.13–1.71] | 0.002 | 0 | 不适用 | 不适用 | |||||||
| jxjy11225394 | MMP-8 | 江等人 | 亚洲人 | 1184/1211 | 3 | C&T | 杂合子模型 | 固定效果 | 0.63 [0.51–0.77] | < 0.00001 | 21 | AAB(相当于 AAA) | 强的 |
|
jxjy1800629 |
肿瘤坏死因子-α |
彭等人 |
白种人/亚洲人 |
432/760 |
7 |
G&A |
等位基因模型 | 固定效果 | 0.73 [0.57–0.93] | 0.012 | 17.50 | 不适用 | 不适用 |
| 纯合子模型 | 固定效果 | 0.52 [0.26–1.05] | 0.069 | 10.00 | 不适用 | 不适用 | |||||||
| 杂合子模型 | 固定效果 | 0.68 | 0.324 | 6.60 | 不适用 | 不适用 | |||||||
| 显性作用 | 固定效果 | 0.54 [0.27–1.09] | 0.085 | 10.40 | 不适用 | 不适用 | |||||||
| 隐性模型 | 固定效果 | 0.73 [0.54–0.97] | 0.029 | 0 | 不适用 | 不适用 | |||||||
|
jxjy361525 |
肿瘤坏死因子-α |
彭等人 |
白种人/亚洲人 |
432/760 |
3 |
G&A |
等位基因模型 | 固定效果 | 0.27 [0.15–0.49] | < 0.001 | 0 | AAB(相当于 AAA) | 强的 |
| 隐性模型 | 固定效果 | 0.25 [0.14–0.47] | < 0.001 | 0 | AAB(相当于 AAA) | 强的 | |||||||
|
内含子 4 中的 VNTR |
eNOS |
宋和李 |
白种人/亚洲人 |
296/329 |
3 |
4a & 4b |
等位基因模型 | 固定效果 | 3.24 [2.04–5.15] | < 0.001 | 0 | BAB | 中等 |
| 显性作用 | 固定效果 | 3.50 [2.12–5.78] | < 0.001 | 0 | BAB | 中等 | |||||||
| 193/226 | 2 | 4a & 4b | 隐性模型 | 固定效果 | 4.76 [0.77–59.61] | 0.094 | 0 | 不适用 | 不适用 | ||||
| jxjy1799983 | eNOS | 宋和李 | 亚洲人 | 403/661 | 3 | T&G | 等位基因模型 | 随机效应 | 1.72 [0.80–3.70] | 0.006 | 80.20 | ABB | 中等 |
| jxjy2070744 | eNOS | 宋和李 | 亚洲/北美 | 145/378 | 2 | C&T | 等位基因模型 | 随机效应 | 1.03 [0.19–5.96] | 0.001 | 90 | ABB | 中等 |
|
jxjy2010963 |
血管内皮生长因子 |
洪等人 |
亚洲人 |
697/877 |
3 |
C&G |
纯合子模型 | 固定效果 | 0.82 [0.72–0.93] | 0.002 | 0 | AAB(相当于 AAA) | 强的 |
| 显性作用 | 固定效果 | 0.79 [0.67–0.92] | 0.003 | 0 | AAB(相当于 AAA) | 强的 | |||||||
| 隐性模型 | 固定效果 | 1.29 [1.06–1.59] | 0.015 | 0 | 不适用 | 不适用 | |||||||
| jxjy6025 | 因子 V | 尚等人 | 混合 | 481/867 | 7 | A&G | 纯合子模型 | 固定效果 | 4.55 [2.75–7.52] | < 0.001 | 0 | AAB(相当于 AAA) | 强的 |
粗体表示 OR、p值和遗传方差的异质性,p值 < 0.05
证据分为强、中和弱。当每个标准都获得 A 时,证据被归类为强。当在每个标准中都没有得分 C 但没有 AAA 时,证据被归类为中等。弱证据在三项标准中的一项中记录为 C
在类固醇诱导的股骨头坏死中,ApoB 具有基因突变 jxjy693 和 jxjy1042031,主效应具有显着的p值。特别是,jxjy1042031 在显性遗传模型中显示出非常显着的结果,不同种族的优势比为 2.90,亚洲人的优势比为 4.81。在 Zhou 等人的研究中,ABCB1 jxjy1045642 基因变异在亚洲人中也显示出显着的结果。PAI-1 jxjy1799889 基因突变基因检测在纯合子模型和显性模型中显示出非常显着的p值低于 0.0005,在龚等人的研究中具有中等证据等级。
表3:来自类固醇诱导的股骨头坏死的合格荟萃分析的基因突变基因检测
| 基因突变 | 基因(或近基因) | 参考(数据收集年份) | 种族 | 样本量(病例/对照) | 纳入研究 | 次要等位基因和主要等位基因 | 遗传模型 | 型号类型 | 报告的 OR (95% CI) | 遗传主效应的p值 | 异质性 (%) | 威尼斯标准 | 证据等级 |
|
jxjy693 |
载脂蛋白B |
陈等人 |
混合 |
570(总计) |
4 |
C&T |
等位基因模型 | 不适用 | 2.63 [0.92–7.53] | 0.072 | 58.00 | 不适用 | 不适用 |
| 杂合模型 | 不适用 | 2.46 [1.27–4.77] | 0.008 | 54.50 | 不适用 | 不适用 | |||||||
| 纯合模型 | 不适用 | 7.70 [1.23–48.18] | 0.029 | 24.40 | 不适用 | 不适用 | |||||||
| 隐性模型 | 不适用 | 7.16 [1.19–43.05] | 0.031 | 32.10 | 不适用 | 不适用 | |||||||
| jxjy693 | 载脂蛋白B | 陈等人 | 混合 | 725(总计) | 5 | C&T | 显性作用 | 不适用 | 2.99 [1.71–5.22] | < 0.001 | 31.30 | BBB | 中等 |
| jxjy1042031 | 载脂蛋白B | 陈等人 | 混合 | 572(总) | 4 | G &A/C | 显性作用 | 不适用 | 2.90 [1.49–5.65] | 0.002 | 50.30 | 商业银行 | 虚弱的 |
| jxjy1042031 | 载脂蛋白B | 陈等人 | 中国 | 415(总计) | 3 | G &A/C | 显性作用 | 不适用 | 4.81 [2.05–11.30] | < 0.001 | 0 | BAB | 中等 |
|
jxjy1801133 |
MTHFR |
陈等人 |
混合 |
251(总计) |
3 |
C&T |
等位基因 | 不适用 | 0.92 [0.59–1.44] | 0.710 | 12.90 | 不适用 | 不适用 |
| 杂合模型 | 不适用 | 0.62 [0.33–1.17] | 0.144 | 22.10 | 不适用 | 不适用 | |||||||
| 纯合模型 | 不适用 | 1.24 [0.48–3.21] | 0.653 | 0 | 不适用 | 不适用 | |||||||
| 隐性模型 | 不适用 | 1.54 [0.63–3.76] | 0.339 | 0 | 不适用 | 不适用 | |||||||
| 507(总计) | 4 | C&T | 显性作用 | 不适用 | 0.94 [0.61–1.45] | 0.775 | 23.40 | 不适用 | 不适用 | ||||
|
jxjy1045642 |
ABCB1 |
周等人 |
亚洲/混合 |
336/712 |
7 |
T&C |
等位基因 | 固定效果 | 0.68 [0.54–0.84] | 0.295 | 18.30 | 不适用 | 不适用 |
| 杂合模型 | 固定效果 | 0.73 [0.53–1.00] | 0.722 | 0 | 不适用 | 不适用 | |||||||
| 纯合子模型 | 随机效应 | 0.43 [0.26–0.69] | 0.049 | 58.10 | 不适用 | 不适用 | |||||||
| 隐性模型 | 随机效应 | 0.52 [0.34–0.81] | 0.009 | 70.30 | 不适用 | 不适用 | |||||||
| 显性作用 | 固定效果 | 0.64 [0.48–0.87] | 0.830 | 0 | 不适用 | 不适用 | |||||||
|
jxjy2032582 |
ABCB1 |
周等人 |
混合 |
275/574 |
5 |
T/A &G |
等位基因 | 固定效果 | 0.73 [0.58–0.90] | 0.587 | 0 | 不适用 | 不适用 |
| 杂合模型 | 固定效果 | 0.66 [0.45–0.96] | 0.985 | 0 | 不适用 | 不适用 | |||||||
| 纯合子模型 | 固定效果 | 0.52 [0.34–0.82] | 0.560 | 0 | 不适用 | 不适用 | |||||||
| 隐性模型 | 固定效果 | 0.71 [0.49–1.01] | 0.385 | 3.9 | 不适用 | 不适用 | |||||||
| 显性作用 | 固定效果 | 0.61 [0.43–0.87] | 0.959 | 0 | 不适用 | 不适用 | |||||||
|
jxjy1799889 |
PAI-1 |
龚等人 |
混合 |
108/917 |
4 |
4G/5G |
等位基因模型 | 不适用 | 1.93 [1.15–3.26] | 0.014 | 0.0004 (P) | 不适用 | 不适用 |
| 纯合子模型 | 不适用 | 3.22 [1.67–6.21] | < 0.0005 | 0.00111 (P) | ACB | 中等 | |||||||
| 显性作用 | 不适用 | 2.31 [1.53–3.49] | < 0.0005 | 0.06 (P) | ACB | 中等 | |||||||
| 隐性模型 | 不适用 | 2.30 [1.24–4.30] | 0.009 | 0.162 (P) | 不适用 | 不适用 |
粗体表示 OR、p值和遗传方差的异质性,p值 < 0.05
证据分为强、中和弱。当每个标准都获得 A 时,证据被归类为强。当在每个标准中都没有得分 C 但没有 AAA 时,证据被归类为中等。弱证据在三项标准中的一项中记录为 C
符合条件的荟萃分析的定性方法评估
使用 AMSTAR 工具对研究结果进行定性方法评估,如表中所述表4. 宋等人显示最低分数为 7,而 Yu Zhang 等人最高分10.5。
表 4:使用 AMSTAR 工具对符合条件的荟萃分析进行定性方法学评估
|
参考 |
清单 |
全部的 |
|||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | ||
| 陈等人 | 1 | 0.5 | 1 | 0.5 | 1 | 1 | 0 | 0.5 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 9.5 |
| 江等人 | 1 | 0.5 | 1 | 0.5 | 1 | 1 | 0 | 0.5 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 8.5 |
| 彭等人 | 1 | 0.5 | 1 | 0.5 | 1 | 1 | 0 | 0.5 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 9.5 |
| 宋等人 | 1 | 0.5 | 1 | 0 | 0 | 0 | 0 | 0.5 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 7 |
| 洪等人 | 1 | 0.5 | 1 | 0.5 | 1 | 1 | 0 | 0.5 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 9.5 |
| 周等人 | 1 | 0.5 | 1 | 0 | 1 | 1 | 0 | 0.5 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 9 |
| 尚等人 | 1 | 0.5 | 1 | 0.5 | 1 | 1 | 0 | 0.5 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 7.5 |
| 龚等人 | 1 | 0.5 | 1 | 0.5 | 1 | 1 | 0 | 0.5 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 8.5 |
缩写:0,无;1、是的;1.5,部分是;AMSTAR,一种评估系统评价的测量工具
股骨头坏死诊断与治疗的国际同行分析
在股骨头坏死的基因解码基因检测研究中,非甾体诱导的股骨头坏死变异基因是 MMP-8(jxjy2012390 和 jxjy11225394)、TNF-α(jxjy1800629 和 jxjy361525)、eNOS(内含子 4 中的 VNTR、jxjy1799983 和 jxjy2070744)、VEGF(jxjy2010963) , 和因子 V (jxjy6025)。其中,MMP8、TNF-α、eNOS和因子V的变异具有较高的显着性,尤其是TNF-α(jxjy361525)和eNOS(内含子4中的VNTR)的变异在两种以上的模型中显示显着性。同时,类固醇诱导的股骨头坏死中具有基因突变基因检测的基因是ApoB(jxjy693和jxjy1042031)、ABCB1(jxjy1045642)和PAI-1(jxjy1799889)。其中,ApoB和PAI-1中包含的基因变异的作用具有较高的显着性,特别是PAI-1(jxjy1799889)中的变体在两种以上的模型具有显着性。
PAI-1 基因是纤溶的重要调节因子,因为它作为纤溶酶原激活物的抑制剂发挥作用。jxjy1799889是PAI-1基因中研究最多的多态性,在675个核苷酸位置有鸟嘌呤缺失。PAI-1-675 4G 等位基因显示出比 PAI-1-675 5G 等位基因更高的转录活性,而 PAI-1-675 4G 等位基因与血浆中更高的 PAI-1 水平有关。在这项研究中,jxjy1799889 的等位基因和隐性模型显示出与类固醇诱导的股骨头坏死高度显着相关。也就是说,PAI-1 4G 变体的携带者发生类固醇诱导的股骨头坏死的风险高于 5G 变体的携带者。似乎4G等位基因是一个重要的致病因素,其基本功能是干扰PAI-1基因的表达。PAI-1基因的低表达可能是纤维蛋白溶解低的股骨头坏死的原因。
MMP-8 通常在中性粒细胞中表达,但也在软骨细胞和滑膜成纤维细胞中表达,位于染色体 11q22.3。MMP-8与股骨头坏死关联的确切机制是佳学基因解码正在进一步深入研究的课题,但多项研究表明,这是对骨组织重塑和发育中细胞外基质分解途径的调节作用的结果,或者MMP-8可能影响股骨头的损伤和炎症。在股骨头坏死的基因解码研究中,jxjy11225394的杂合子模型具有很强的证据等级和显着性,p值 < 0.00001,非类固醇诱导的股骨头坏死的平均 OR 为 0.63。推测MMP-8对非甾体诱导的股骨头坏死具有保护作用。
基因解码明确指出,像 TNF-α 基因 308 或 -238 这样的 TNF-α 启动子多态性会影响转录活性。一些研究表明,非类固醇诱导的股骨头坏死的病理生理学与骨细胞和成骨细胞的凋亡有关。此外,据报道 TNF-α 会影响骨细胞和成骨细胞释放与破骨细胞增殖和成熟相关的细胞因子。因此,TNF-α的多态性可以改变成骨细胞和破骨细胞的活化,从而导致非甾体诱导的股骨头坏死恶化。在股骨头坏死的基因解码研究中,作为 TNF-α 多态性的 jxjy1045642 在等位基因和隐性模型中表现出高度显着性。在等位基因和隐性模型中,平均 OR 分别为 0.27、0.25,这可能意味着 TNF-α 的下调对非类固醇诱导的股骨头坏死具有保护作用。
ABCB1 基因编码转运蛋白 p-糖蛋白 (P-gp),它在将外来物质泵出细胞方面具有关键功能。在Ning Han等的研究中,BMSCs成骨细胞成熟的前体,由于 BMSCs 减少,当P-gp在类固醇诱导的股骨头坏死中被抑制时,发现骨髓间充质干/基质细胞(BMSC)的过度脂肪生成,表明股骨头中的骨形成受到抑制。在这项研究中,类固醇诱导的股骨头坏死中的 jxjy1045642 降低了具有次要 T 等位基因的疾病发病风险。说明ABCB1基因可能是调节激素诱导的股骨头坏死的重要基因。
已知位于染色体 7q35-q36 上的 eNOS 基因会增加 NO。已知 NO 介导血管生成和血管舒张。然而,它的关键作用是预防血栓形成,这是非类固醇诱导的股骨头坏死的主要病理生理机制。内含子 4 中的 27 bp 可变 VNTR 是 eNOS 的多态性,包括野生 4b 等位基因和突变的 4a 等位基因。这种 4b/a 多态性与 NO 血浆浓度降低有关。在这项研究中,eNOS 基因中的 4b/a 多态性在非甾体诱导的股骨头坏死中显示出高度的意义。这可能意味着eNOS的表达降低可能导致非类固醇诱导的股骨头坏死的发生。此外,eNOS 基因中的 jxjy1799983 和 jxjy2070744 多态性也显示出显着性,但它们的异质性太高(jxjy1799983 为 80.20%,jxjy2070744 为 90%)。
已知 VEGF 在股骨头坏死的坏死区高度表达。它的主要作用是血管生成和成骨。还报道了 VEGF 基因促进骨髓细胞和近内皮细胞增殖。在这项研究中,被称为 VEGF -634G/C 多态性的 jxjy2010963 具有很高的显着性。对所有遗传模型进行评估时,纯合子和显性模型的平均 OR 低于 1.00,而隐性模型的平均 OR 超过 1.00,表明 jxjy2010963 的遗传模型可能在预测股骨头坏死风险方面具有重要作用。
因子 V (FV) 是众所周知的参与血液凝固过程的成分。因子 V Leiden (FVL) 基因突变是 FV 的典型多态性,可导致血栓形成倾向和血管内凝血障碍。多项研究表明,FVL 基因突变引起的血栓形成可能是非类固醇诱导的股骨头坏死的主要病因。在股骨头坏死的基因解码研究中,作为FVL基因多态性的jxjy6025在纯合子模型中表现出高度显着性。平均 OR 为 4.55。有强有力的证据。这可能意味着因子 V 的下调可能会增加股骨头坏死发生的风险。
APO 是通过与细胞受体相互作用介导正常脂质代谢的主要血浆蛋白。据报道,APO 是评估类固醇诱导的股骨头坏死中脂质代谢紊乱的敏感标志物。在 Karami 等人的一项研究中,jxjy693 的 C 等位基因与高家族胆固醇水平高度相关。相比之下,T 等位基因的存在具有保护作用。jxjy1042031 也被认为对调节 ApoB 与胆固醇受体的结合能力很重要。在股骨头坏死的基因解码研究中,作为 ApoB 多态性的 jxjy693 和 jxjy1042031 在优势模型中均表现出高显着性。他们的平均 OR 超过 2.5,这意味着如果 ApoB 功能不佳,股骨头坏死发生率的风险可能会增加。这些结果表明,脂质代谢紊乱可能导致股骨头脂肪生成,这是类固醇诱导的股骨头坏死的主要原因之一。
简而言之,非类固醇诱导的相关基因是MMP-8、TNF-α、eNOS、VEGF和因子V,类固醇诱导的相关基因是ApoB、ABCB1和PAI-1。股骨头坏死的总体基因机制如图 1 所示。
相关的整体基因机制.jpg)
图 2:与股骨头坏死(FHON)相关的整体基因机制。红色箭头表示股骨头坏死的危险因素,绿色箭头表示股骨头坏死的保护因素
有待继续进步的股骨头坏死基因解码基因检测
股骨头坏死的基因解码研究有几个局限性。首先,股骨头坏死的基因解码基因检测重大课题研究组的研究在数据收集中排除了性别因素。其次,没有具体说明种族的分类。第三,未对研究结果进行进一步验证。第四,虽然这项研究包括了来自各种论文的许多 SNP 结果,但它受到当时可用研究的限制。第五,股骨头坏死的基因解码研究包含大量来自各种参考文献的荟萃分析。因此,结果可能导致高估一个或另一个基因的实际意义。最后,荟萃分析中报告的大多数基因突变基因检测仅在搜索包含基因突变的荟萃分析时出现一次,以仅收集与类固醇诱导的股骨头坏死和非类固醇诱导的股骨头坏死相关的可靠数据。
股骨头坏死基因解码基因检测结论
基于对已发表论文的系统回顾,股骨头坏死的基因解码基因检测重大课题研究组组织了与股骨头坏死相关的基因组并检查了每个贡献。股骨头坏死的基因解码基因检测重大课题研究组的结果可以为理解股骨头坏死病因机制提供一种综合方法。预计这些结果可能有助于预诊断策略和评估非类固醇诱导和类固醇诱导的股骨头坏死的个体风险。
更多与本文内容相关的科学依据:
Integrative analyses of genes related to femoral head osteonecrosis: an umbrella review of systematic reviews and meta-analyses of observational studies.
Published online 2022 Mar 28. doi: 10.1186/s13018-022-03079-4
J Orthop Surg Res。2022; 17:182
